
Photo of actual Lyrodesma Hinge Teeth. They are preserved in calcite.
Put forth as one of many possible explanations to the mystery fossil by Roger Cuffey of Penn State,
"The interlocking teeth on the hinge of pelecypod ("bivalve") shells. Several of your 3- & 4-plate specimens look a lot like some of those, such as on trigoniids & heterodonts. If so, these would likely be molds/impressions/casts (of those structures), because most (though not all) pelecypods' shells are aragonite, and so would have dissolved away or inverted to calcite long ago."
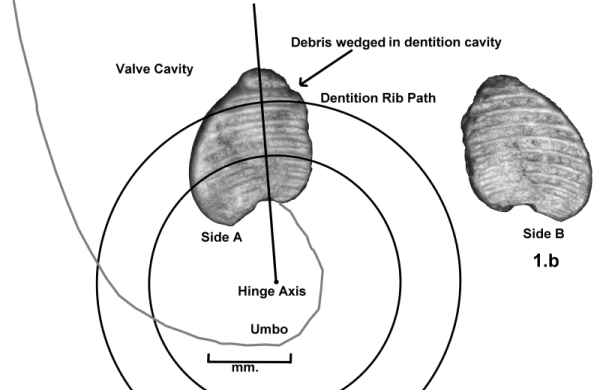
I began looking for specimens of pelecypods from the type-Cincinnatian that have sufficient shell preservation to show details of their hinge teeth. I found some specimens of Lyrodesma (species undetermined) from the Kope Formation, Southgate member of Carrollton, Kentucky that had shell preservation. Among them were two specimens that showed the hinge teeth. On one specimen you can see that there are ridges between the teeth. These ridges tell me that Lyrodesma teeth could have formed these mystery plates as molds between them.
This specimen has matrix down in there between the teeth, so I'm still looking for a specimen that can tell me if the ridges are curved in the same direction as is evident on the multi-plate specimens of the mystery fossil.
Here are some images of the Lyrodesma from the "Southgate" site in Carrollton Kentucky.
Photo of actual Lyrodesma Hinge Teeth. They are preserved in calcite.

The specimen is a single valve filled with matrix consisting of mostly crinoid
material. The hinge teeth as seen at the top of the picture.

Here is a closeup of the hinge teeth showing the ridges on the sides of the
teeth. Note the orientation of the teeth.
See more photos of Lyrodesma hinge dentitions preserved
in calcite.
See photos of "Mystery Fossil"
multi-element specimens for comparison.
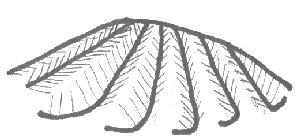
Each tooth is convex and terminates at relatively sharp edges. The mystery plates, on the other hand, terminate with a blunt, smooth surface. The above hand drawing illustrates the edges of the hinge teeth.
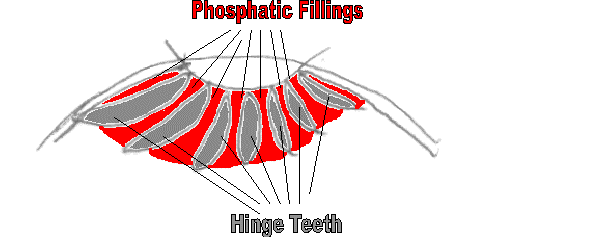
If the mystery plates were fillings between the teeth, they should be concave,
which we do find. The mystery plates have what appears to be a rough,
"open" edge (
see this illustration
), and a smooth, "closed" edge as well as a generally wedged shape.
This may be consistent with pelecypod teeth external molds if the smooth,
"closed" edge of the molds were facing the back of the pelecypod valve
(making contact with the valve surface) and the rough "open" edge were
facing the open end of the valve. This is mostly conjecture, as I need to find a
more complete specimen of Lyrodesma that shows the hinge teeth all the
way back to the valve surface with no matrix obscuring the view.

The
multi-plate specimens
of the mystery fossil resemble the Lyrodesma teeth in that they are
oriented in radial patterns in a similar way to the pelecypod teeth. Most of the
multi-plate mystery fossils form a 90 degree angle with four plates. Some,
however, form as little as a 45 degree angle with four plates.

Here is another view of the Lyrodesma hinge teeth, this time looking
down from the umbo and with greater contrast to show the ridges on the sides of
the teeth.
In checking out the evidence I have found that ridged impressions were common in this Cyclora Fossil Hash and that the "nodules" found hanging out of the sides of the mystery fossils have been found to be both gastropod steinkerns and crinoid columnals. That page is titled Secondary Phosphatization .
Once I was convinced that my "Mystery Fossil" was Lyrodesma hinge dentition molds, Rich Fuchs, a Dry Dredger who helps organize the Cincinnati Museum collections at the Geier Collections and Research Center, told me he had found an example of my Mystery Fossil that had been identified in 1972 By Bill Harrison as Lyrodesma hinge teeth. Click here to read about this amazing discovery.
As a result of this and subsequent samples I gathered and measured, I have written the following description for these phosphatic molds of Lyrodesma sp. hinge dentitions. I am hoping it is more professionally written than what you have read in this pages before this point.
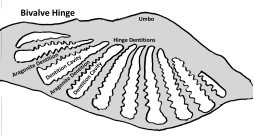
PHOSPHATIC MOLDS OF BIVALVE HINGE DENTIONS IN THE PHOSPHATIC LAYERS OF
THE CINCINNATIAN SERIES
William P Heimbrock, Dry Dredgers
Bivalves smaller than 2 mm., which constitute less than 5
percent of the fauna in the phosphatic layers I sampled, are represented as phosphatic steinkerns. Larger bivalves
are preserved only with phosphates in intergranular porosity between the hinge
dentitions. Numerous examples of Lyrodesma sp. dentitions were found with the
surface features preserved as a thin phosphate film less than 0.1 millimeters
thick. In no case was the area between dentitions completely filled by this
film. Instead, the area between the
Lyrodesma sp. dentitions were filled with bits and pieces of other
phosphatic fossils, especially gastropod steinkerns and echinoderm stereom.
The occurrence of these Lyrodesma sp. dentition molds
constitutes between 0.5 and 1.0 percent of the phosphatic fossils at the
Florence Freedom site as well as other exposures of these layers in the C4
sequence. The available
number of dentition mold specimens and their multiphasic development, provide us
with another excellent platform for understanding multiphasic intergranular
porosity.
These dentition molds were first identified in 1972 by
William Harrison while examining a specimen with four elements in the Cincinnati
Museum of Natural History collection (CNMH
IP70662) which was labeled as collected by a Mr. Jenkins in 1966. The
specimen was found in the Fairmount formation of the edrioasteroid site of
Forestville, Ohio (North end of excavation for Beechmont Mall shopping center,
opposite Anderson High School. Ohio coordinate system: 397,300’; 1,474,700’
Bell, 1976 pp. 88, 121, 163) with the description “Problematical radiate ‘multituberculate’
structures”. William Harrison
amended the label for the specimen to indicate “Lyrodesma
dentition”.
In the year 2000, this author (Heimbrock) found countless
thousands of these dentition molds on the aforementioned Florence Freedom
Ballpark site. After years of
examining the single and multi-element specimens and sending samples to
professional colleagues around the globe, the diagnosis of Lyrodesma sp.
dentition molds was confirmed.
The genera Lyrodesma was first described in 1841 by T. A.
Conrad in the Fifth annual report on the palaeontology of the State of New York:
New York Geol. Survey Ann. Rept. 5, p. 25-57.
Replacement of aragonite shell material with calcite in
bivalves Cycloconcha sp. and Lyrodesma sp. are common in the Carrollton beds of
the Southgate member of the Latonia (Kope) formation in Northern Kentucky.
However, in zones of low carbonate sedimentation, such as the phosphatic
micromorph layers of the Cincinnatian C4 Sequence, the original aragonite clam
valves were not replaced with calcite. Gritty hash consisting of phosphatic
steinkerns of mollusks and bits of phosphatic material from intergranular
porosity with echinoderm stereom and bryozoan zooecia wedged between aragonite
hinge teeth. The wedged grit permitted phosphates to form a film on the surfaces
of the hinge dentitions while it was still preserved in aragonite. The
secondarily phosphatized film and phosphate grit were sturdy and retained the
cavities between bivalve hinge dentitions as long as the ribbed surface features
after aragonite deterioration.
Reworking may have occurred after the decay of the
aragonite hinge, causing the dentition molds to disarticulate from the original
radial orientation of the dentitions. Most of these phosphatic dentition molds
are found as individual plates and some fragment and become part of the gritty
infill for intergranular porosity
with other shell cavities in the hash.
Process of Phosphogenesis of Bivalve Hinge Dentition Molds
During a long period of sediment starvation, shell material
was subject to subtidal winnowing.
Individual valves of pelecypods were partially degraded, leaving primarily the
thickest part of the valve – the hinge. In other cases, most or all of the valve
may have remained complete until eventual burial in a sandy material consisting
largely of phophates. A mold of a
hinge dentition could not have formed if the two valves remained articulated.
There was no cavity in that case to receive phosphates or phosphatic debris. The
gaps between the dentitions of a single valve provided the optimal size for tiny
fragments of material that has already undergone phosphatic porosity.
It seems likely that these odd bits of ribbed dentition
molds formed in phases of intergranular porosity, as illustrated in the
illustration sequence below.
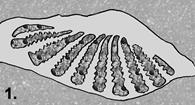
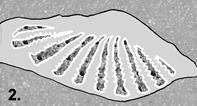

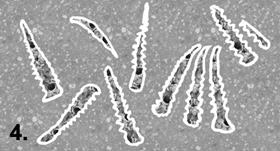
Step 1: Burial.
Gritty hash consisting of phosphatic steinkerns of mollusks and bits of
phosphatic material from intergranular porosity with echinoderm stereom and
bryozoan zooecia wedged between aragonite hinge teeth.
Step 2:
A phosphatic film is built up between the gritty debris and the
dentitions of the aragonite valve.
Step 3: Over time the aragonite valve dissipates, leaving the phosphatic
film molding the external surfaces and ribbing of the dentitions and encasing
the grit between the dentitions. The individual dentition molds are now
unattached from one another and free to disassociate with reworking of the
matrix.
Step 4: Reworking can
occur after the decay of the aragonite hinge, causing the dentition molds to
separate rom the original radial orientation and become individual elements
encased in phosphatic film with the negative impression of the ribbed surface of
an individual dentition.
Identification of association between phosphatic molds and
Lyrodesma hinge dentitions was made first by finding articulated examples of
phosphatic dentition molds with multiple dentitions still in the original radial
orientation. In this preservation, the resemblance to hinge dentitions is
apparent. Individual elements are more common and can be identified as Lyrodesma
hinge dentition molds by the following criteria.
1.
By possessing a phosphatic film on all but one side of the element.
2.
By having 4 to 11 ribs embossed on two opposite
sides of the element.
3.
By being filled with miscellaneous and random
bits of material composed mostly of phosphates. This material may be a
combination of echinoderm stereom, mollusk steinkerns, bryozoan zooecia infils
and other grains smaller than the space between hinge dentitions.
4.
By having a length usually about 1.5 millimeters
with examples found up to 2.25 mm.
5.
By having a width of about half the length.
6.
By having a depth that varies depending on the
side.
7.
By having ribs that are concentrically curved.
All ribs on both sides of the element have an arc with a consistent axis.
8.
By having a longest edge that extends and
connects the phosphatic film from either side. (Molds the crotch of the hinge
between dentitions).
9.
By having next longest edge that is open and
exposes the miscellaneous phosphatic grit filling the interior of the element.
10.
By having a narrowest edge that is filled with
the phosphatic film and is the thickest point on the element. This edge is
always the farthest point (apex) from the axis circumvented by the ribs.
11.
By having
a thinnest edge circumvented by the concentric ribs indicating the closest point
to the axis.
12.
By having ribs on both sides that appear to be
closer together the closer they are to the apparent axis.
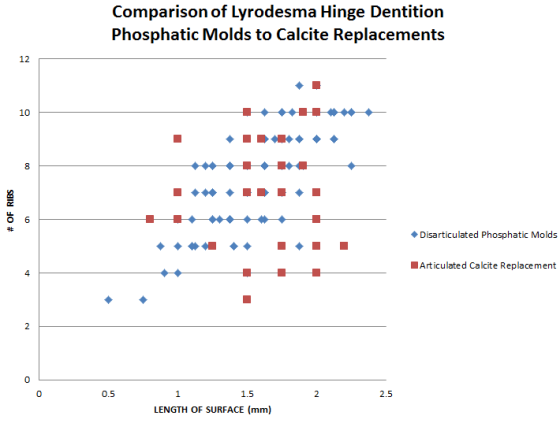
Scattergram 1:
A sample of complete valves of Lyrodesma preserved in calcite was taken from the
Carrollton beds (Kope Fm) of Carrollton KY and Covington KY. These were measured
and dentition ribbing, when present, counted. This was compared to the
disarticulated phosphatic molds of bivalve dentitions found on the Florence KY
Arnheim Fm. (C4 Sequence) Phosphatic layer. The analysis shows that the sizes of
the phosphatic dentition molds was consistent with that of adult Lyrodesma from
the Carrollton beds. The number of ribs acts as a control to indicate short,
fragmented or poorly preserved dentitions.
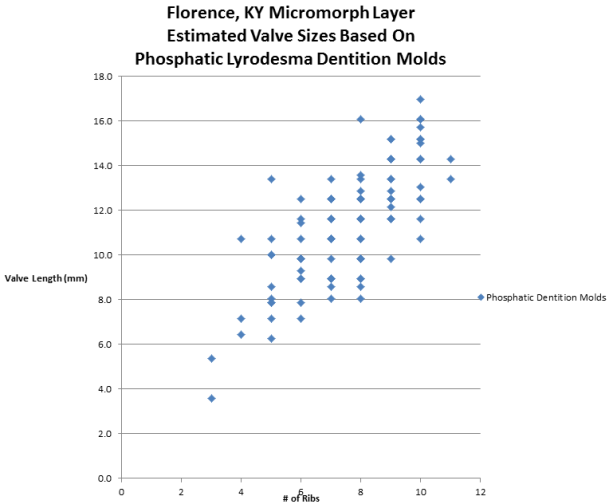
Scattergram 2:
The same sampling of phosphatic dentition molds were measured along the open
edge (the wide edge with debris visible) and used to estimate the total valve
length. From examining the Calcite replacements of complete Lyrodesma valves
from the Carrollton beds (Kope Fm.) of Carrollton, KY and Covington, KY, a ratio
of 7.14 to 1 (X 0.14) between the valve length and the dentition lengths was
found to be the average across all of the dentitions. The cavities between the
calcite dentitions was measured rather than the calcite dentitions themselves so
as to compare correctly to the phosphatic molds. The number of ribs act as a
control in identifying short, fragmented or poorly preserved phosphatic
dentition molds. The result shows that the phosphatic molds represented valves
from 6 to 16 mm in length. If 10 ribs per dentition represents a complete
dentition, then the original valves ranged from 11 to 17 mm.
Click here for a description of the measurements and methods I used.
Bill Heimbrock
1621 Westwood Ave
Cincinnati, OH 45214
(513) 921-2335
billheim@cinci.rr.com
{kind=link}